
There is a specific patch of the human brain, tucked along the upper edge of the temporal lobe, that lights up when we hear a person speak. For years, neuroscientists have called these the temporal voice areas, or TVA. The standard assumption was that this region is a specialized piece of biological hardware, a “conspecific” processor tuned exclusively to the sounds of our own kind. It was a neat, tidy boundary between us and the rest of the animal kingdom.
But biology is rarely that tidy. New research from a team at the University of Geneva suggests that this neural real estate is less of a private club and more of an ancestral inheritance. When we hear the hoot of a chimpanzee, our temporal voice areas react with a selective intensity that was previously thought to be reserved only for Homo sapiens.
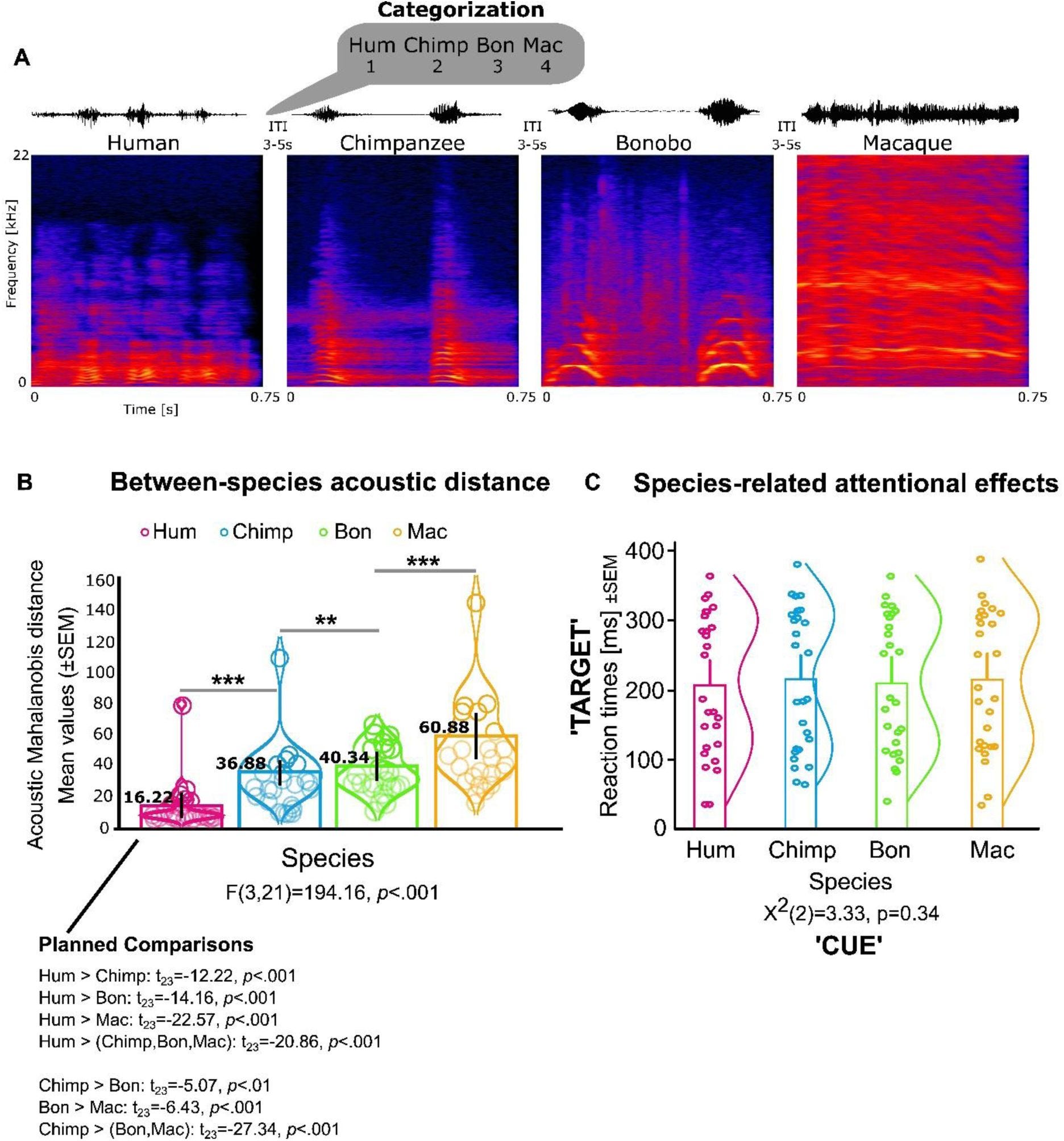
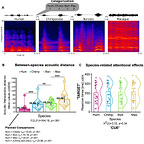
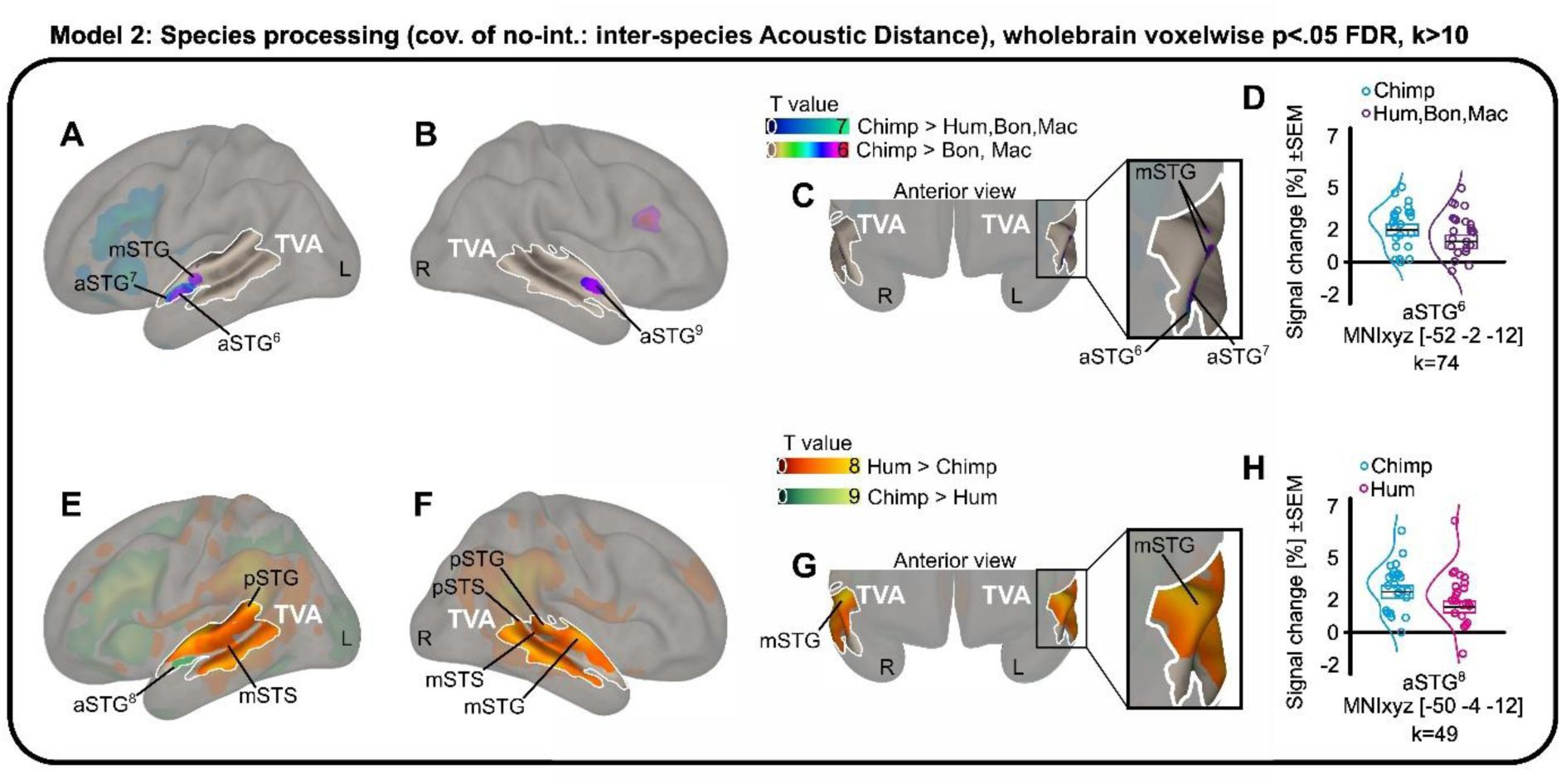
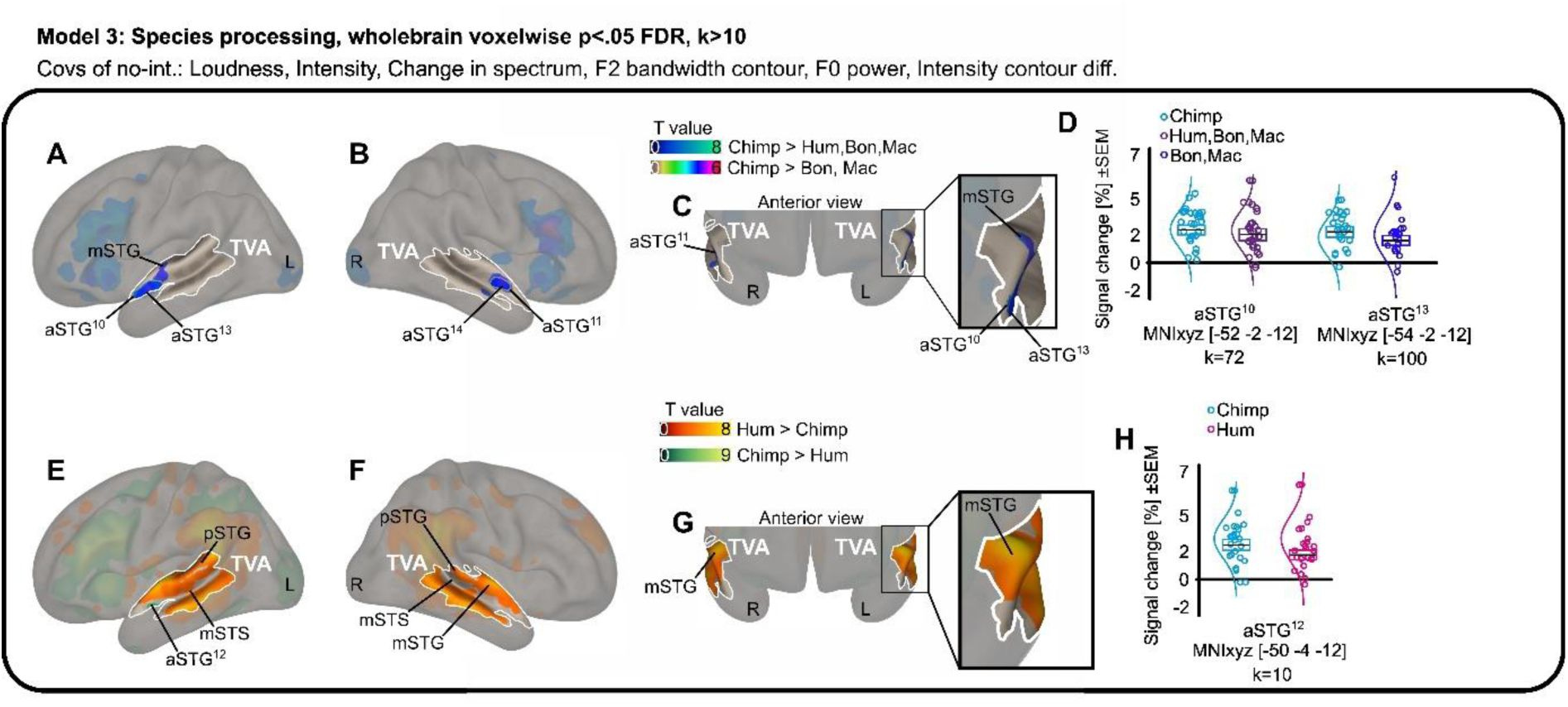
This isn’t just a general reaction to noise. The researchers1 used fMRI to watch the brains of 23 participants as they listened to a variety of primate calls: humans, rhesus macaques, bonobos, and chimpanzees. Even after the team mathematically stripped away the basic acoustic differences like pitch and loudness, a specific subregion in the anterior superior temporal gyrus remained stubbornly active for chimpanzee vocalizations.
It appears that our “human” voice areas are actually tuned to a broader frequency range that includes the sounds of our closest evolutionary neighbors. Interestingly, this neural kinship did not extend to bonobos. Despite being just as closely related to us as chimpanzees, bonobo calls failed to trigger the same specialized response in the human TVA.
The Acoustic Gap
The reason for this distinction likely lies in the anatomy of the throat. Bonobos have a shorter larynx, which gives their vocalizations a much higher fundamental frequency than those of humans or chimpanzees. Chimpanzee voices, by contrast, sit much closer to our own acoustic “sweet spot”.
The researchers hypothesized that the human auditory cortex evolved a tonotopic map tailored to the specific frequencies of human speech. Because chimpanzee calls fall within or near that range, our brains process them using the same specialized machinery we use for our parents, friends, and strangers on the street. Bonobos, through a process of evolutionary neoteny—the retention of juvenile traits into adulthood—seem to have drifted away from that shared acoustic space. Their calls are simply too high-pitched for our voice-processing hardware to recognize them as “voice-like”.
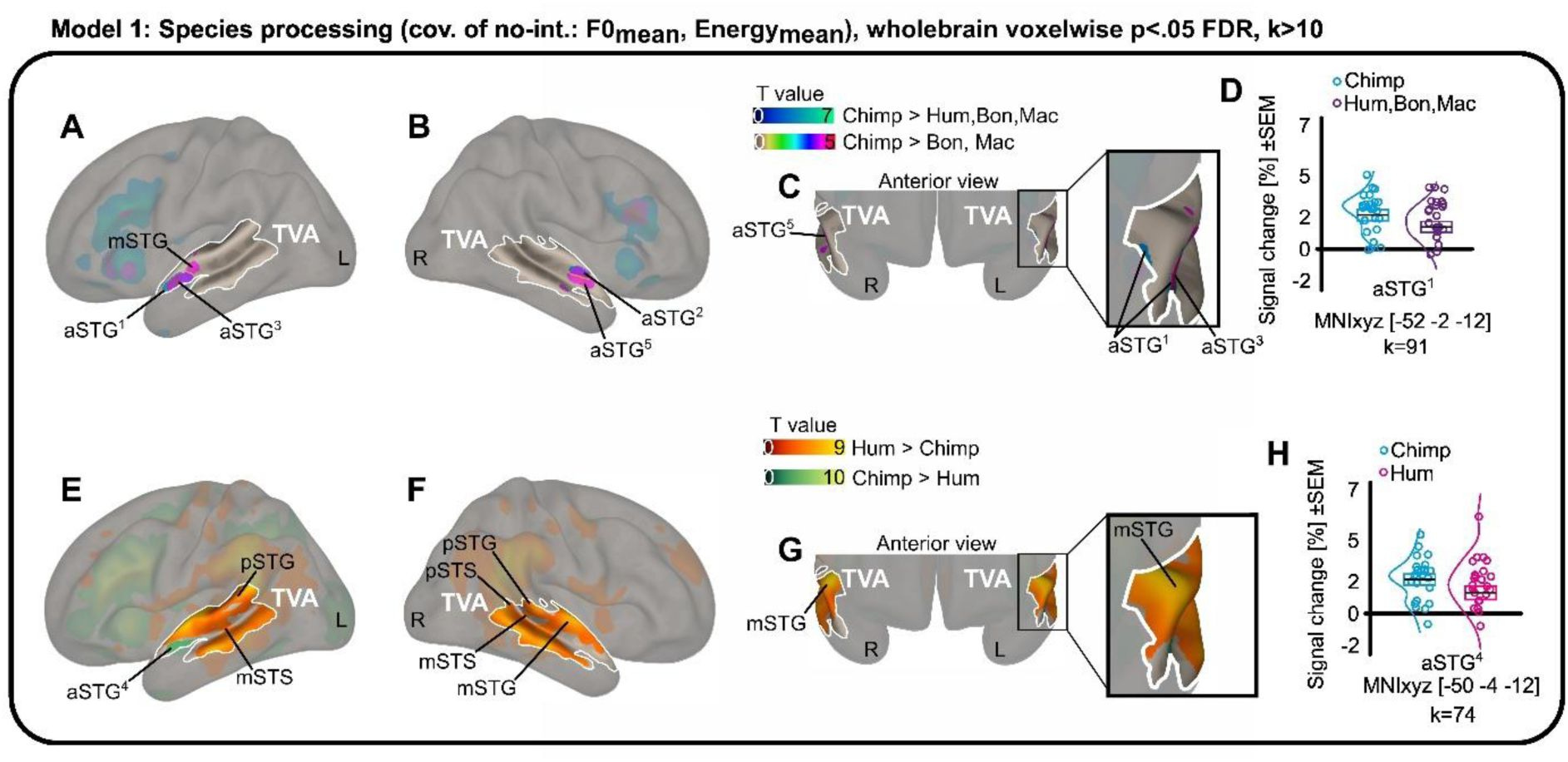
This finding challenges the idea of human “uniqueness” in brain organization. If our voice-sensitive areas are also chimpanzee-sensitive, it suggests that these regions didn’t appear out of nowhere to support human language. Instead, they likely represent an ancient system for social monitoring that we share with the other great apes. We didn’t build a new room for the human voice; we just redecorated an existing one.
Evolutionary Echoes
This leaves us with a compelling question about the nature of our own perception. When we hear a chimpanzee, are we hearing an “animal,” or is our brain instinctively categorizing that sound as a kind of distorted, familiar “other”?
The data showed that while the human voice still generates the most widespread activity across the temporal cortex, the anterior subregions of the TVA are remarkably sensitive to these cross-species similarities. It is as if the back of the brain handles the complex nuances of human identity and emotion, while the front remains vigilant for the basic, structural signatures of a primate throat.
We often think of our brains as being uniquely adapted to the modern world, but studies like this remind us that we carry deep, prehistoric echoes in our anatomy. Our “human” brain is still, in many ways, a primate brain, reaching out across six million years of separation to recognize the sound of a cousin.
Ceravolo, L., Debracque, C., Gruber, T., & Grandjean, D. (2025). Sensitivity of the human temporal voice areas to nonhuman primate vocalizations. eLife.







